
Respirazione Aerobica
FOSFORILAZIONE OSSIDATIVA
Il ciclo di Krebs e la glicolisi producono una minima parte di ATP rispetto alla resa finale della respirazione aerobia. Come viene prodotta altra energia? La risposta si trova nella riossidazione del NADH e del FADH2.
Il NADH e il FADH2 sono riossidati grazie alla fosforilazione ossidativa (fosforilazione perché fosforilo ATP, lo formo; ossidativa perché utilizzo l’ossigeno), un processo che si attua mediante la catena di trasporto degli elettroni sulla membrana interna del mitocondrio.
LA CATENA DI TRASPORTO DEGLI ELETTRONI
Nel processo di fosforilazione ossidativa gli elettroni ad alta energia sono trasferiti dal NADH e dal FADH2 all’ossigeno per mezzo della catena respiratoria, producendo ATP per un meccanismo chemiosmotico.
La catena di trasporto degli elettroni nella membrana interna del mitocondrio, comprende quattro complessi e trasportatori di membrana. I complessi sono proteine trans membrana, ovvero enzimi che ricevono elettroni dai coenzimi e li trasportano fino all’accettore cioè l’ossigeno con il quale formano acqua nella matrice. I trasportatori di membrana invece sono proteine “navetta” che trasportano gli elettroni da un complesso all’altro riducendosi e riossidandosi.
TRASPORTO E- ATTRAVERSO I COMPLESSI
Andando con ordine analizziamo ogni complesso e ogni trasportatore che intervengono nella riossidazione di NADH. I due elettroni vengono ceduti al
Complesso I, chiamato anche complesso NADH deidrogenasi, contenente gruppi prostetici ferro- zolfo. Ciascun atomo di ferro si alterna tra lo stato ossidato (Fe3+) e quello ridotto (Fe2+). I due elettroni vengono ceduti al trasportatore, il coenzima q meglio noto come ubichinone, risiedente nella porzione interna della membrana. L’ubichinone trasporta gli elettroni al Complesso III chiamato complesso del citocromo b/c1, il quale li accetta e li cede al trasportatore sulla parte esterna della membrana interna, noto come citoctromo c. Quest’ultimo contiene anch’esso il ferro che si riduce e ossida, ma all’interno di un gruppo prostetico eme. Il citocromo c (che trasporta un elettrone alla volta) cede gli elettroni al Complesso IV, chiamato complesso della citocromo ossidasi contenente un gruppo prostetico contenente rame e ferro che si ossidano e riducono. Questo termina col cedere gli e all’ossigeno formando acqua.
IL COMPLESSO II E IL FADH2
Quando invece si tratta di FADH2 entra in gioco il complesso II chiamato anche succinil-coenima Q ossidoreduttasi. Questo si trova nella parte interna della membrana. gli elettroni del FADH2 non sono ceduti al complesso I ma direttamente al II che li trasporta all’ubichinone, per poi continuare come avviene per il
NADH. Ciò che differenzia il complesso II dagli altri è il fatto che non si tratta di una pompa protonica (quindi non pompa protoni). Questo comporta che vengano pompati 2 protoni in meno, motivo per il quale vedremo la resa del FADH2 produce meno ATP.
GRADIENTE PROTONICO
Quando gli elettroni passano nei complessi liberano energia permettendo il pompaggio di protoni H+ dalla matrice allo spazio intramembrana generando un pH acido. I complessi I III IV dunque funzionano anche da pompe protoniche e creano un gradiente protonico: il complesso I al passaggio di 2e pompa 4H+, il complesso III al passaggio di 2e pompa 4H+ e il complesso IV al passaggio di 2e pompa 2H+. Il totale di protoni pompati al passaggio di due protoni è 10H+. Il gradiente protonico genera una forza motrice protonica che è la somma della variazione di pH e del potenziale d’azione (dato dal potenziale di membrana). Questa non è la sola teoria proposta ma è la più accreditata. Venne scoperta da Mitchell nel 1961 e prende il nome di teoria chemiosmotica.
ATP SINTASI
La caratteristica essenziale del modello chemiosmotico è il legame fra il trasporto degli elettroni e la sitesi di ATP è potenziale elettrochimico attraverso una membrana. Abbiamo detto che con il pompaggio di protoni si viene a creare una forza motrice protonica, ma come si forma l’ATP? Grazie all’ATPsintasi.
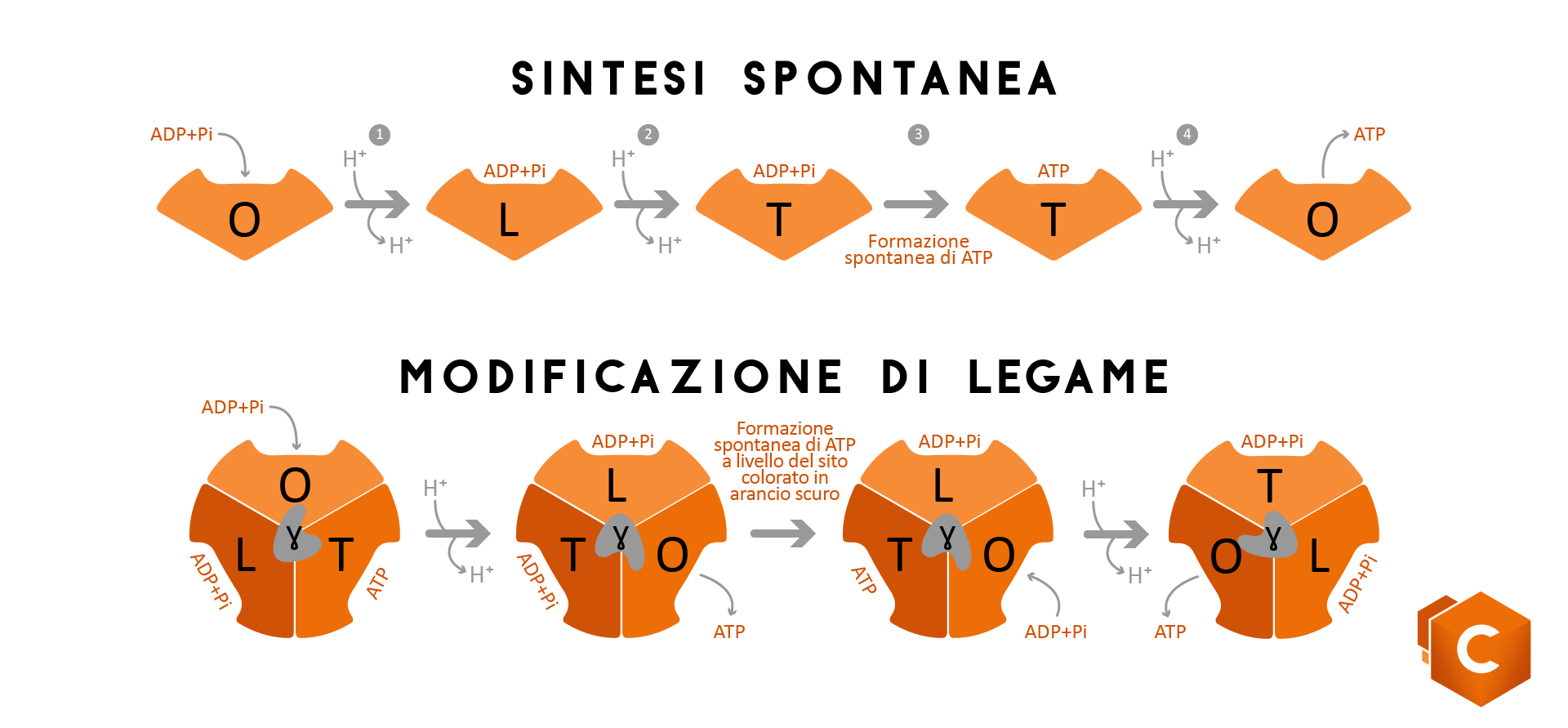
L’ATPsintasi è un complesso multiproteico formato da due subunità F0e F1uniti da un sottile stelo. La subunità F0, si trova inserita nel doppio strato fosfolipidico della membrana interna del mitocondrio e forma un canale nel quale passano i protoni spinti dal gradiente, mentre la subunità F1 si trova nella matrice mitocondriale.La subunità F0, all’entrata di un protone spinto dal gradiente, ruota e lo rilascia alla subunità F1 che utilizza l’energia libera prodottasi dal movimento del protone che va nella matrice per avvicinare i reagenti ovvero ADP e P. Esistono due teorie che spiegano la formazione di ATP:
- una teoria, quella della sintesi spontanea, prevede che la subunità F1 possa assumere 3 conformazioni al passaggio di ogni rispettivo protone: una conformazione (O)nella quale ADP+ P stanno per entrare, una conformazione (L) nella quale sono i substrati sono leggermente legati e una terza conformazione (T) nella quale sono legati e hanno formato ATP. A questo punto altri protoni entrano e la produzione ricomincia.
- Un’altra teoria invece prevede che la sintesi di ATP avvenga per modificazione di legame e che il sito F1 sia formato da 3 facce catalitiche. Secondo questa teoria al passaggio di protoni avviene un cambiamento simultaneo di tutti e tre i siti e la produzione di ATP ogni volta in un sito diverso.
In casi rari se la concentrazione degli H+ è maggiore nella matrice l’ATPsintasi funziona al contrario.
Dal momento che per ogni molecola di NADH vengono pompati circa 10 protoni, e che per formare ATP servano 3-4 protoni, si conclude che per ogni NADH si ricavano circa 3 molecole di ATP.
Mentre, per quanto riguarda FADH2, (dal momento che entra nel secondo complesso e che questo non pompa H) per ogni molecola vengono pompati meno protoni quindi la resa in ATP è di 2.
Comments: 2
E’ errata la formula del piruvato
Che svista😱 grazie mille di averci avvisato, provvediamo subito a correggere!